Mechanisms of Meiosis – MOM
Katja WASSMANN


Sexual reproduction requires the generation of haploid gametes through two meiotic divisions, named meiosis I and II. In mammals and especially humans, female gametogenesis is highly error prone, leading to the generation of oocytes harbouring the wrong chromosome content.
Projects in my group aim at dissecting the molecular mechanisms underlying the two meiotic divisions with a focus on oocytes, to understand how errors occur. The central question we study deals with the issue of how meiosis I-specific events are executed only in meiosis I, and those of meiosis II only in meiosis II.
Keywords: Meiosis, Oocytes, Cohesin, Phosphoproteome, Cell cycle, Aneuploidy
+33 (0)157278148 katja.wassmann(at)ijm.fr https://www.researchgate.net/profile/Katja-Wassmann
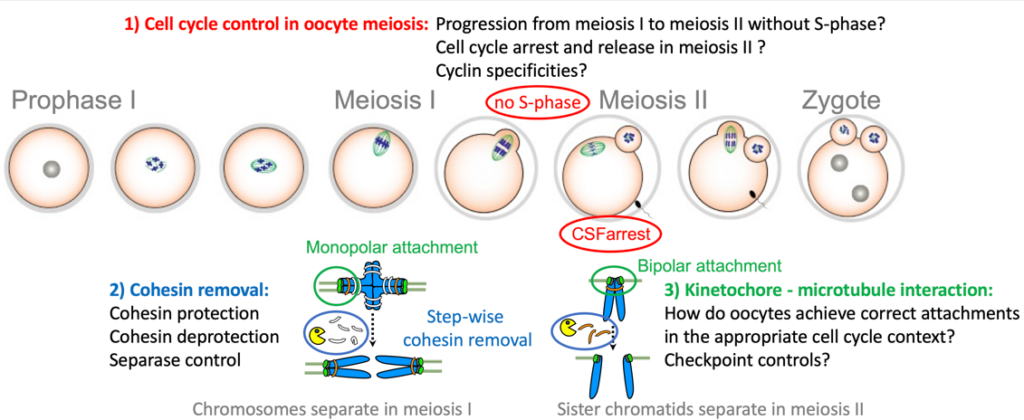
- In meiosis, key events need to happen in the correct order, and at the right time, otherwise the generation of healthy gametes is compromised. In our projects we focus on elucidating the underlying regulatory mechanisms of meiosis-specific events with a special focus on oocytes, to gain insights into how these events are put into place in the correct developmental context, e.g. meiosis I or II.Specific features of the meiotic divisions (meiosis I and II):
- Homologous chromosomes (each consisting of two sister chromatids) are paired and segregated in meiosis I, and sister chromatids in meiosis II
- Kinetochores (the attachment sites for the bipolar spindle) of the two paired sister chromatids are co-oriented to the same pole in meiosis I, and bioriented to opposite poles in meiosis II (named mono- and bipolar orientation or attachment, respectively).
- The cohesin complex, which is holding sister chromatids together, is removed in a step-wise manner, from chromosome arms in meiosis I, and the centromere region in meiosis II.
- Transition from meiosis I into meiosis II occurs without intervening S-phase
An additional specificity of vertebrate oocytes is the requirement to implement two cell cycle arrests: One before entry into meiosis I, in prophase I, which lasts decades in humans; a second arrest in metaphase of meiosis II, called CSF-arrest, to await fertilization.
The overarching question of our projects is the following:
How are the two successive meiotic divisions implemented without mixing up meiosis I and meiosis II specific events?
Our research questions:

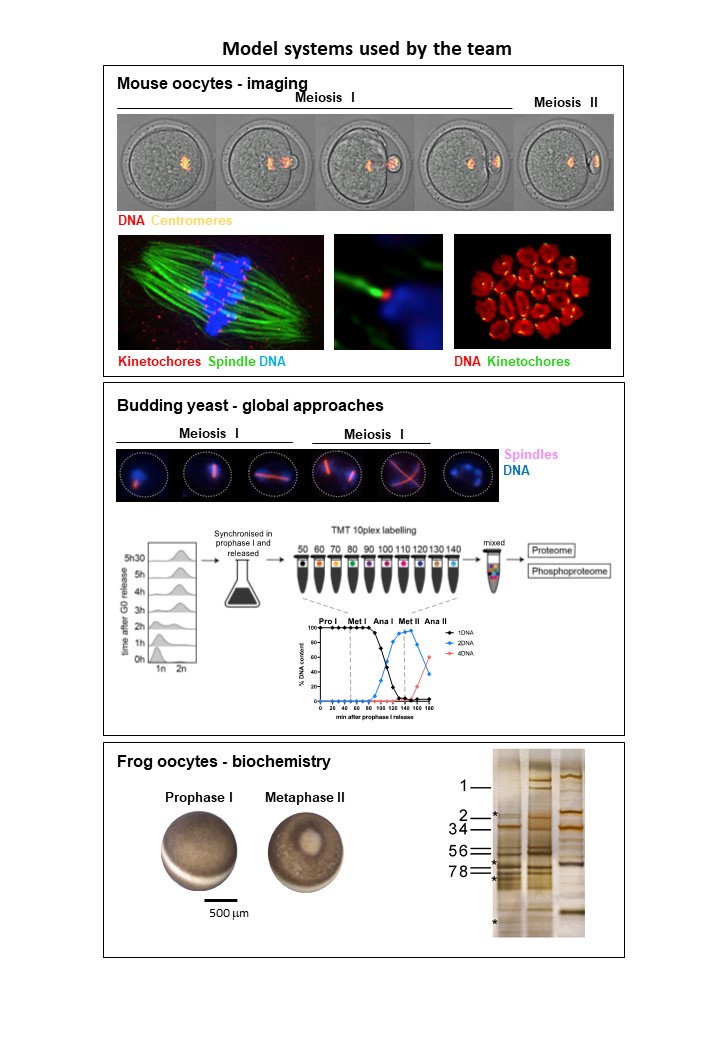
- Model systems we use in the lab:
2) Step-wise cohesin removal in mammalian oocytes
Step-wise cohesin removal is brought about by protection of the centromeric cohesin subunit Rec8 from cleavage by the protease Separase at the centromere region in meiosis I, and by removing this protection before sister chromatid segregation in meiosis II. Protection is due to the recruitment of Sgo2, which brings the phosphatase PP2A to the centromere to maintain Rec8 there in a dephosphorylated state. However, several open questions are remaining, starting with the question of how Separase activity is controlled to get inhibited and activated twice, and how proteins involved in protection do so in meiosis I, but not meiosis II, and only in the centromere region.
3) Kinetochore attachments and checkpoint controls in mammalian oocytes
Again related to our main question of how meiosis I and meiosis II events are implemented at the correct time, we want to understand how checkpoints can recognize the correct attachments in their proper context, e.g. monopolar attachments to segregate chromosomes in meiosis I, and bipolar attachments in meiosis II to segregate sister chromatids. Additionally we address whether both kinetochores are functional in meiosis I when they are mono-oriented and fused.

Group Leader:
Katja WASSMANN
Phone : +33 (0)157278148
Mail : katja.wassmann (at) ijm.fr
Members:
Eulalie BUFFIN, MCF
Aude DUPRE, CR
Sandra TOUATI, CR
Safia EL JAILANI, PhD Student
Damien CLADIERE, Engineer
Samih EL AARAJ, Master 2
Emmanuelle MARJAULT, Engineer
Irem POLAT, PhD Student


Bouftas N.*, Schneider L.*, Halder M., Demmig R., Baack M., Cladière D., Walter M., Al Abdallah H., Kleinhempel C., Messaritaki R., Müller J., Passarelli F., Wehrle P., Heim A., Wassmann K.#+ and Mayer T.U.#
Cyclin-B3 prevents Emi2/Xerp1 from setting up precocious CSF-arrest in oocyte meiosis I,
Dev. Cell (2022) 57, 2305-2320
(*co first-authors, #co-corresponding authors, +lead author)
Nikalayevich E., El Jailani S., Cladière D., Gryaznova Y., Fosse C., Touati S.A., Buffin E., and Wassmann K.
Aurora B/C-dependent phosphorylation promotes Rec8 cleavage in mammalian oocytes
Current Biology (2022) doi: 10.1016/j.cub.2022.03.041, in press
Gryaznova Y., Keating L.*, Touati S.A.*, Caldière D., El Yakoubi W., Buffin E., and Wassmann K.
Kinetochore individualization in meiosis I is required for centromeric cohesin removal in meiosis II,
EMBO J. (2021) 40, e106797 (*co second-authors)
Mehmet E. Karasu*, Bouftas N.*, Keeney S., and Wassmann K.
Cyclin B3 promotes anaphase I onset in oocyte meiosis,
J Cell Biol. (2019) 218, 1265-1281 (*co first-authors)
Vallot A., Leontiou I., El Yakoubi W., Cladière D., Bolte S., Buffin E.*, and Wassmann K.*+
Tension-induced error correction, and not kinetochore attachment status activates the SAC in an Aurora-B/C-dependent manner in oocytes,
Current Biology (2018) 28, 130-139 (*co-corresponding authors, + lead author )
El Yakoubi W., Buffin E., Cladière D, Gryaznova Y., Berenguer I., Touati S.A., Gómez R. , Suja J.A.,van Deursen J.M., and Wassmann, K.
Mps1 kinase-dependent Sgo2 centromere localization mediates cohesin protection in mouse oocyte meiosis I,
Nature Communications (2017) DOI: 10.1038/s41467-017-00774-3
Touati S., Buffin E., Cladière D., Rachez C., Hached K., van Deursen J.M., and Wassmann K.
Mouse oocytes depend on BubR1 for proper chromosome segregation but not prophase I arrest,
Nature Communications (2015) DOI: 10.1038/ncomms7946
Chambon J.P., Touati S., Berneau S., Cladière D., Hebras C., Groeme R., McDougall A., and Wassmann K.
The PP2A inhibitor I2PP2A is essential for sister chromatid segregation in meiosis II,
Current Biology (2013) 23, 485-490
Touati S., Cladière D, Lister L.M., Leontiou I., Chambon J.P., Rattani A., Böttger F., Stemmann O., Nasmyth K., Herbert M., and Wassmann K.
Cyclin A2 is required for sister chromatid segregation, but not Separase control, in mouse oocyte meiosis,
Cell Reports (2012) 29, 1077-1087
Kudo N.R.*, Wassmann K.*, Anger M., Schuh M., Wirth G.K., Xu H., Helmhart W., Kudo H., Mckay M., Maro B., Ellenberg J., de Boer P., and Nasmyth K.
Resolution of Chiasmata in Oocytes Requires Separase-Mediated Proteolysis,
Cell (2006) 126, 135-146 (*co first-authors)
Wassmann K.*, Niault T., and Maro B.
Metaphase I Arrest Upon Activation of the Mad2-Dependent Spindle Checkpoint in Mouse Oocytes
Curr. Biol. (2003) 13, 1596-1608 (*corresponding author)
Publications
Reviews
Book chapters
Thesis defended:
- Khaled Hached: 2006-2010
- Sandra Touati: 2011- 2014
- Antoine Vallot: 2014-2017
- Nora Bouftas: 2015-2019
- Leonor Keating: 2017-2021
Thesis in progress:
- Antoine Langeoire-Cassalta: 2019-
- Dunja Čelebić: 2020-
- Safia El Jailani: 2021-
Thomas Mayer, University of Konstanz, Germany
Evelyn Houliston, Laboratoire Océanologique de Villefranche-sur-Mer, France
Equipe FRM 2021-2024
ANR JCJC 2022-2025
ANR PRC 2019
We are recruiting an engineer (information to follow)